

Extraído del libro, Audiología: Teoría y Práctica. Capítulo 2. Anatomía funcional y fisiopatología de los sistemas auditivo, vestibular y fonador.
El conjunto de las complejas cavidades del interior de la porción petrosa o peñasco del temporal constituye el laberinto óseo del oído interno, con una parte vestibular o del equilibrio (vestíbulo y 3 conductos semicirculares) y otra coclear o auditiva (cóclea o caracol).
LABERINTO ÓSEO Y MEMBRANOSO
El conjunto de las complejas cavidades del interior de la porción petrosa o peñasco del temporal constituye el laberinto óseo del oído interno, con una parte vestibular o del equilibrio (vestíbulo y 3 conductos semicirculares) y otra coclear o auditiva (cóclea o caracol). Estos espacios contienen líquido perilinfático o perilinfa que rodea a sacos y conductos de paredes epiteliales (laberinto membranoso), a su vez rellenos de líquido endolinfático o endolinfa, en cuyas paredes hay zonas especializadas en órganos sensoriales para la audición y el equilibrio. Existe un balance estable entre la producción y eliminación de los líquidos laberínticos para mantener una presión constante en los respectivos espacios.
La endolinfa, semejante al suero sanguíneo por su predominio en iones K+, se produce por secreción activa y trasudación a partir de la sangre que circula por capilares de la estría vascular del conducto coclear y en el plano semilunar de las ampollas de los conductos semicirculares.
El vestíbulo es un espacio ovoideo (4 mm) que comunica con la cavidad timpánica a través de las ventanas oval y redonda.
Gracias a su función metabólica aporta oxígeno, intercambia iones y retira residuos hacia el espacio meníngeo epidural a través del saco endolinfático. La perilinfa, semejante al líquido cefalorraquídeo (LCR) por su predominio en iones Na+, procede de un trasudado de los vasos laberínticos y del LCR contenido en el espacio subaracnoideo, con el que se comunica a través del conducto perilinfático o acueducto coclear. Su función es protectora y de sostén del laberinto membranoso. La cortilinfa es similar a la perilinfa por su riqueza en iones Na+ y está contenida en los pequeños espacios de Nüel y túnel de Corti delimitados por las células del órgano de Corti. Estos quedan aislados de la rampa media con endolinfa y de la timpánica con perilinfa (Suárez y cols., 2007).
El vestíbulo es un espacio ovoideo (4 mm) que comunica con la cavidad timpánica a través de las ventanas oval y redonda. Los conductos perilinfático y endolinfático (o acueducto del vestíbulo) comunican el vestíbulo con la cavidad craneal, la cual, así mismo, comunica a través del conducto auditivo interno (CAI) con la base de la cóclea, donde se localiza una lámina ósea con numerosos orificios (criba espiroidea) atravesados por las fibras del nervio coclear que, junto con la porción vestibular del VIII par craneal o nervio vestibulococlear o estatoacústico, penetran en la región del bulbo raquídeo. Por el CAI también pasa el nervio facial (VII par).
El conjunto de las complejas cavidades del interior de la porción petrosa o peñasco del temporal constituye el laberinto óseo del oído interno, con una parte vestibular o del equilibrio (vestíbulo y 3 conductos semicirculares) y otra coclear o auditiva (cóclea o caracol).
Al vestíbulo también tienen entrada y salida los tres conductos semicirculares (CS) óseos (2/3 de círculo), los cuales presentan una dilatación o ampolla en uno de sus extremos y están orientados en los tres planos del espacio. Uno en el plano horizontal, el CS lateral o externo, y dos en planos verticales: el superior en el plano sagital y el posterior en el plano frontal.
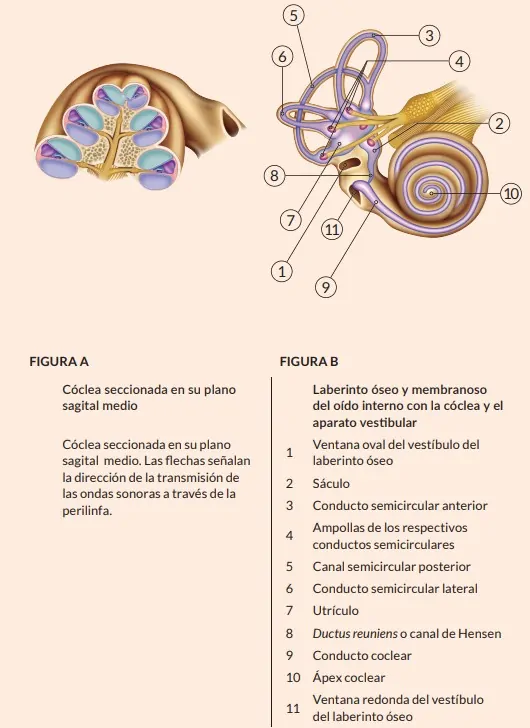
Del conjunto de estructuras epiteliales que forman el laberinto membranoso, las correspondientes al aparato vestibular son el sáculo, el utrículo (en el interior del vestíbulo) y los conductos semicirculares membranosos, situados en el interior de sus respectivos espacios óseos.
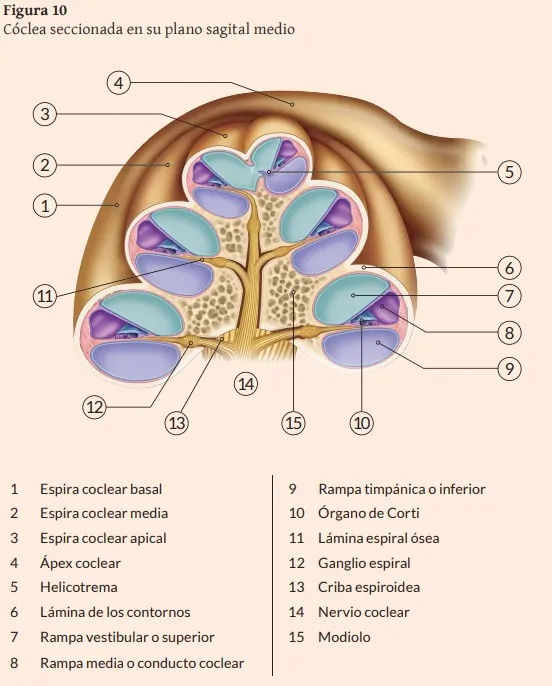
Los conductos superior y posterior confluyen en un único conducto común que desemboca en el vestíbulo. Desde el vestíbulo parte el conducto espiral óseo de la cóclea, con un fondo ciego, describe dos vueltas y tres cuartos en el humano (35 mm de longitud total) alrededor de un eje central óseo hueco de forma cónica llamado modiolo o columela. Su diámetro (2-3 mm) va disminuyendo progresivamente desde la base hacia el ápex o vértice, terminando en fondo cerrado. Toda la cóclea tiene unos 5 mm de altura y unos 9 mm de anchura en la base (Figuras 1,2,4,8,9) (Angulo y cols., 1997; Gil-Carcedo, 2011; Suárez y cols., 2007; Williams, 1998). Dicho conducto espiral óseo está dividido parcialmente en dos rampas: vestibular o superior y timpánica o inferior. Están separadas por una delgada lámina espiral ósea que parte del modiolo, de forma similar al relieve espiral de un tornillo. Coincidiendo con el origen de la lámina espiral ósea, en el modiolo se encuentra el canal espiral de Rosenthal, que aloja a las neuronas del ganglio espiral de Corti, cuyos axones son las fibras del nervio coclear que se dirige al CAI y sus dendritas atraviesan la lámina espiral ósea, hasta contactar con la base de las células sensoriales o ciliadas externas e internas del órgano de Corti.
Del conjunto de estructuras epiteliales que forman el laberinto membranoso, las correspondientes al aparato vestibular son el sáculo, el utrículo (en el interior del vestíbulo) y los conductos semicirculares membranosos, situados en el interior de sus respectivos espacios óseos.
El conducto coclear membranoso corresponde al aparato auditivo y se extiende desde el vestíbulo, donde comunica con el sáculo por el conducto reuniens o de Hensen, al vértice de la cóclea ósea. En una sección transversal al eje del conducto coclear, este tiene un perfil triangular. Ocupa una región intermedia, denominada rampa media, entre la lámina espiral ósea y la lámina de los con- tornos o pared externa de dicho conducto espiral óseo. Contiene endolinfa y las estructuras celulares del órgano sensorial auditivo u órgano de Corti (Figura 10).
La membrana de Reissner (dos capas de células epiteliales aplanadas) constituye la pared superior o techo del conducto coclear. Formando un ángulo agudo con el suelo, va desde la lámina espiral ósea hasta la pared externa del con- ducto espiral. Limita con el espacio de la rampa vestibular o superior, la cual se relaciona con la cavidad timpánica a través de la ventana oval del vestíbulo (ocupada por la platina del estribo y su ligamento anular).
El hecho físico por el que se originan las ondas viajeras se fundamenta en que anatómicamente la membrana basilar va disminuyendo su grosor y aumentando su anchura desde la base hacia el ápex.
La membrana basilar está en la pared inferior o suelo del conducto coclear y sobre ella se encuentra el órgano de Corti. Limita con la rampa timpánica o inferior, la cual se relaciona con la cavidad timpánica a través de la venta- na redonda cerrada por el tímpano secundario. A lo largo de todo el conducto coclear, la membrana basilar se sitúa entre el borde libre de la lámina espiral ósea y el ligamento espiral externo, por el que se fija a la pared externa de la cóclea. Esta distancia aumenta progresivamente desde las porciones más basales hacia las más apicales de la cóclea. De forma inversa, el grosor de la membrana basilar va disminuyendo desde basal hacia apical, a medida que los diámetros de las fibras de colágeno que la constituyen se van adelgazando.
A lo largo de todo el conducto coclear, la membrana basilar se sitúa entre el borde libre de la lámina espiral ósea y el ligamento espiral externo, por el que se fija a la pared externa de la cóclea.
La estría vascular está formada por un epitelio con vasos sanguíneos, a partir de los cuales se produce la endolinfa por filtrado activo. Se localiza en la pared lateral del conducto coclear, adherida a la superficie ósea del conducto espiral. En el ápex coclear se encuentra el helicotrema o punto de conexión entre las rampas vestibular y timpánica ocupadas por perilinfa, que comunica con la del vestíbulo y la del resto de espacios perilinfáticos (Figura 10) (Angulo y cols., 1997; Gil-Carcedo, 2011; Suárez y cols., 2007; Williams, 1998)
ÓRGANO SENSORIAL DE LA AUDICIÓN
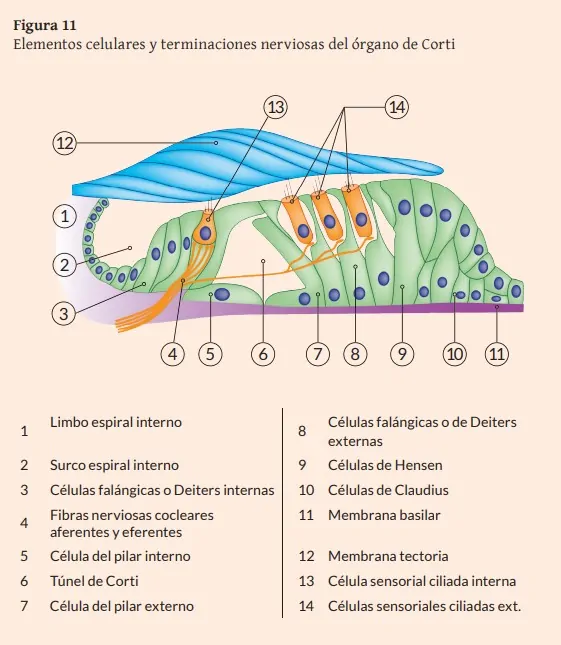
El órgano de Corti u órgano sensorial de la audición es de origen neuroectodérmico. Sus células son de tipo epitelial especializado y están diferenciadas según su morfología y función en células de soporte y células sensoriales (con características similares a las neuronas ya que sintetizan neurotransmisores que liberan en sus contactos sinápticos). Se disponen principalmente sobre la membrana basilar del conducto coclear y está parcialmente cubierto por la membrana tectoria. La configuración global de este agregado celular varía ligeramente desde las espiras basales hacia las apicales de la Cóclea (Figura 11) (Suárez y cols., 2007).
La estría vascular está formada por un epitelio con vasos sanguíneos, a partir de los cuales se produce la endolinfa por filtrado activo. Se localiza en la pared lateral del conducto coclear, adherida a la superficie ósea del conducto espiral.
Existen dos tipos de células sensoriales o receptoras según su localización en el órgano de Corti en relación al eje central del modiolo coclear. Las células ciliadas internas se disponen formando una fila a lo largo del conducto coclear. Están situadas por encima de las células de soporte falángicas internas. En su superficie apical hay tres hileras de cilios en línea recta, con un tamaño decreciente, y unidos entre sí por puentes filamentosos. Las células ciliadas externas se disponen en tres filas sobre las células falángicas externas. Sus cilios son similares a los de las internas, pero distribuidos en forma de V. Los extremos de los cilios se relacionan con la superficie inferior de la membrana tectoria situada sobre ellos, existiendo un mayor anclaje a ella a medida que las células ciliadas son más externas. La membrana tectoria, paralela a la membrana basilar, es más estrecha en la base de la cóclea y alcanza doble anchura en las regiones más apicales. Consta de una porción medial delgada, anclada firmemente al limbo espiral, y una porción lateral libre más gruesa, que se sitúa sobre el órgano de Corti. La membrana tectoria es acelular y está compuesta por colágeno, glicoproteínas y glucosaminoglicanos (formando filamentos inmersos en una matriz amorfa mucopolisacárida) que son sintetizados por las células interdentales del limbo espiral interno situado sobre la lámina espiral ósea. Entre el limbo y las células falángicas internas se encuentra el surco espiral interno tapizado por un epitelio de células cúbicas.
Uno de los tipos de células de soporte son las células de los pilares externo e interno (con forma piramidal alargada y abundantes microtúbulos en su citoesqueleto que le confiere gran consistencia) que configuran el túnel de Corti. Este espacio de sección triangular contiene cortilinfa y se encuentra central en el órgano de Corti. Por su interior pasan fibras nerviosas aferentes (prolongaciones dendríticas de las neuronas del ganglio espiral) y eferentes que se dirigen a la base de las células sensoriales ciliadas externas e internas.
Otras de las células de soporte son las de Deiters o falángicas externas e internas, cuyas prolongaciones digitiformes se ensanchan en el extremo y se unen lateralmente a la región apical de las respectivas células ciliadas externas e internas situadas sobre ellas y bañadas por la cortilinfa que ocupa los espacios de Nüel. Este mosaico de células sensoriales y de soporte unidas por desmosomas constituye la lámina o membrana reticular, la cual delimita la rampa media con perilinfa, de los espacios con cortilinfa del órgano de Corti.
Las células de soporte situadas lateralmente a las células de Deiters externas son las células de Hensen, con forma alargada y las células de Claudius, más cortas y que tapizan el surco espiral externo (Figura 11) (Angulo y cols., 1997; Gil-Carcedo, 2011; Rodríguez y Smith-Agreda, 2004; Suárez y cols., 2007; Williams, 1998).
FUNCIÓN DEL OÍDO INTERNO
Dada la naturaleza mecánica de las ondas de presión sonora que son transmitidas al oído interno, bien por vía aérea (a través de las estructuras del oído externo y medio), o bien por vía ósea (a través de las vibraciones de los huesos del cráneo), el órgano sensorial de Corti actúa como un mecanorreceptor, transformando la energía mecánica del estímulo sonoro en impulsos nerviosos bioeléctricos que serán transmitidos a neuronas de la vía aditiva aferente.
Esta específica función de transducción es realizada por las células receptoras ciliadas externas e internas. Además, a nivel coclear se realiza una función tonotópica o cocleotópica, es decir, una primitiva discriminación tonal (discernir entre frecuencias de sonidos presentados consecutivamente) según la región de la cóclea estimulada.

Existen dos tipos de células sensoriales o receptoras según su localización en el órgano de Corti en relación al eje central del modiolo coclear. Las células ciliadas internas se disponen formando una fila a lo largo del conducto coclear.
Esto se explica por dos hechos: por un lado, la estimulación de un grupo de células sensoriales situadas sobre una determinada zona de la membrana basilar («teoría del lugar») que se desplaza en sintonía con la frecuencia del sonido, ya que sus características morfológicas le confieren las propiedades físicas adecuadas para ello, por otro, la específica, compleja y coordinada inervación aferente (excitatoria) y eferente (inhibitoria) de las células sensoriales, hace posible una sintonización o selección frecuencial del sonido por contraste entre dos zonas muy próximas de células ciliadas estimuladas (Haines, 2013; Pickles,1988; Purves, 2015; Suárez y cols., 2007).
Dada la naturaleza mecánica de las ondas de presión sonora que son transmitidas al oído interno, bien por vía aérea, o bien por vía ósea, el órgano sensorial de Corti actúa como un mecanorreceptor, transformando la energía mecánica del estímulo sonoro en impulsos nerviosos bioeléctricos que serán transmitidos a neuronas de la vía aditiva aferente.
Los mecanismos esenciales del proceso de transducción de las células ciliadas del órgano de Corti se fundamentan en cambios en los potenciales endococleares (voltajes de los líquidos intra y extracelulares de la cóclea). Ello es consecuencia de la apertura de canales iónicos provocada por microdeformaciones en la membrana citoplasmática de los cilios de las células sensoriales (unidos entre sí por puentes filamentosos), cuando son desplazados o curvados lateralmente por los movimientos radiales de la membrana tectoria sobre la membrana reticular (Pickles,1988; Suárez y cols., 2007). Estos desplazamientos relativos entre las dos membranas paralelas son originados por las oscilaciones de las ondas sonoras, transmitidas por los líquidos laberínticos.
Los mecanismos esenciales del proceso de transducción de las células ciliadas del órgano de Corti se fundamentan en cambios en los potenciales endococleares (voltajes de los líquidos intra y extracelulares de la cóclea).
Según la intensidad y frecuencia del sonido, tanto en la audición por vía aérea, como en la audición por vía, se generan oscilaciones en la perilinfa que se distribuyen por la rampa vestibular de la cóclea hasta el helicotrema en microsegundos.
Según la intensidad y frecuencia del sonido, tanto en la audición por vía aérea (cuando la presión sonora se transmite desde el oído externo y medio al oído interno a través de la base del estribo que se desplaza en la ventana oval), como en la audición por vía ósea (cuando las vibraciones sonoras se transmiten por los huesos del cráneo directamente al oído interno), se generan oscilaciones en la perilinfa que se distribuyen por la rampa vestibular de la cóclea hasta el helicotrema en microsegundos. Luego alcanza el final de la rampa timpánica, donde el tímpano secundario cierra la ventana redonda y compensa dicha presión con desplazamientos alternos a los del estribo. Al mismo tiempo, la presión oscilatoria de la perilinfa actúa sobre las membranas del conducto coclear (de Reissner y basilar con el órgano de Corti) que, a su vez, se transmite a la endolinfa contenida en él y a la membrana tectoria. Esta permanece casi inmóvil por estar unida al limbo espiral interno, por el poco peso específico de sus componentes y por quedar prácticamente anuladas las presiones en sentidos opuestos, transmitidas desde la perilinfa de las rampas vestibular y timpánica a la endolinfa del conducto coclear. Por tanto, los movimientos desencadenados en la membrana basilar son los más importantes en el proceso de audición.
La membrana tectoria, paralela a la membrana basilar, es más estrecha en la base de la cóclea y alcanza doble anchura en las regiones más apicales. Consta de una porción medial delgada, anclada firmemente al limbo espiral, y una porción lateral libre más gruesa, que se sitúa sobre el órgano de Corti.
Por su naturaleza elástica y con cierta inercia, se forman ondas viajeras u ondas de desplazamiento que van siempre desde la base hacia el ápex de la cóclea. La velocidad de transmisión de estas ondas por la membrana basilar es más lenta que por el líquido, tardando unos 2 a 5 milisegundos en recorrer los 35 mm de la cóclea (de 2 vueltas y ¾ en el humano). (Pickles,1988; Purves, 2015; Suárez y cols., 2007).
El hecho físico por el que se originan las ondas viajeras se fundamenta en que anatómicamente la membrana basilar va disminuyendo su grosor y aumentando su anchura desde la base hacia el ápex. De forma inversa, la lámina espiral ósea es más ancha en la base y se va estrechando hacia las espiras apicales. Dado que cerca de la ventana oval la membrana basilar tiene mayor cantidad de masa por unidad de longitud, oscila preferentemente con sonidos de altas frecuencias, ya que en esta zona alcanza su máxima amplitud y entra en resonancia su frecuencia de vibración con la del sonido.
Por el contrario, los sonidos de baja frecuencia desencadenan una onda viajera con una pequeña amplitud en la base que irá aumentando hacia el ápex, a medida que la estructura física de la membrana va cambiando a menor cantidad de masa por longitud. Estas evidencias justifican, según la hipótesis de la «teoría del lugar», la discriminación tonal cocleotópica. Ello consiste en que sonidos de cada frecuencia del campo auditivo harán que un determinado lugar de la membrana basilar alcance su máxima amplitud de oscilación de la onda viajera correspondiente. Así pues, en cada precisa región coclear todas las estructuras del órgano de Corti dispuestas sobre ella se mueven en bloque, ya que las células de los pilares y la lámina reticular aportan rigidez al conjunto celular, y por tanto, los cilios de las células sensoriales reciben una mayor presión mecánica. Además, los desplazamientos relativos de la membrana tectoria respecto a la membrana reticular (en la base de los cilios) son más acentuados en las regiones externas del órgano de Corti que en las internas, existiendo un mayor estímulo mecánico y cizallamiento o incurvación de los cilios de las células ciliadas externas que los de las internas.
A mayor intensidad sonora, mayor número de células ciliadas serán excitadas llevando a cabo su función de transducción a impulso nervioso. (Pickles,1988; Purves, 2015; Suárez y cols., 2007).
Tras el estímulo sonoro ocurre una despolarización o activación de las células ciliadas, que provoca la liberación de un neurotransmisor excitatorio (principalmente glutamato) en su polo basal. Aquí se localizan las sinapsis con las fibras aferentes o dendritas de las neuronas del ganglio espiral de Corti.
En cuanto a los valores de los potenciales endococleares, medidos de forma electrofisiológica en reposo, se sabe que el líquido intracelular de las células del órgano de Corti y las de la estría vascular tienen carga negativa de -70 mV a -80 mV (con escaso K+ y abundante Na+). Se considera que hay un potencial de reposo en la membrana de las células sensoriales cuando las superficies apicales de las células sensoriales y sus cilios están inmersos en endolinfa de carga positiva (+80 mV, con abundante K+) y el resto de la membrana celular por cortilinfa de carga neutra (0 mV, con abundante Na+ y escaso K+, siendo va- lores similares a los de la perilinfa). Las microdeformaciones mecánicas de la membrana celular por el cizallamiento ciliar sincrónico con el estímulo sonoro, tienden a facilitar apertura de canales con corrientes iónicas que cambian el potencial de reposo de las células ciliadas, produciéndose un potencial receptor. Ello es posible gracias a una diferencia de potencial, de al menos 150 mV entre el espacio endolinfático (+80 mV) y el interior de las células ciliadas (-70 mV). Tras el estímulo sonoro ocurre una despolarización o activación de las células ciliadas, que provoca la liberación de un neurotransmisor excitatorio (principalmente glutamato) en su polo basal. Aquí se localizan las sinapsis con las fibras aferentes o dendritas de las neuronas del ganglio espiral de Corti.
En ellas se producen excitaciones sincrónicas que transmiten potenciales de acción a través de los axones del nervio acústico a las neuronas de los núcleos cocleares, segundo nivel de la vía auditiva (Pickles,1988; Suárez y cols., 2007).




