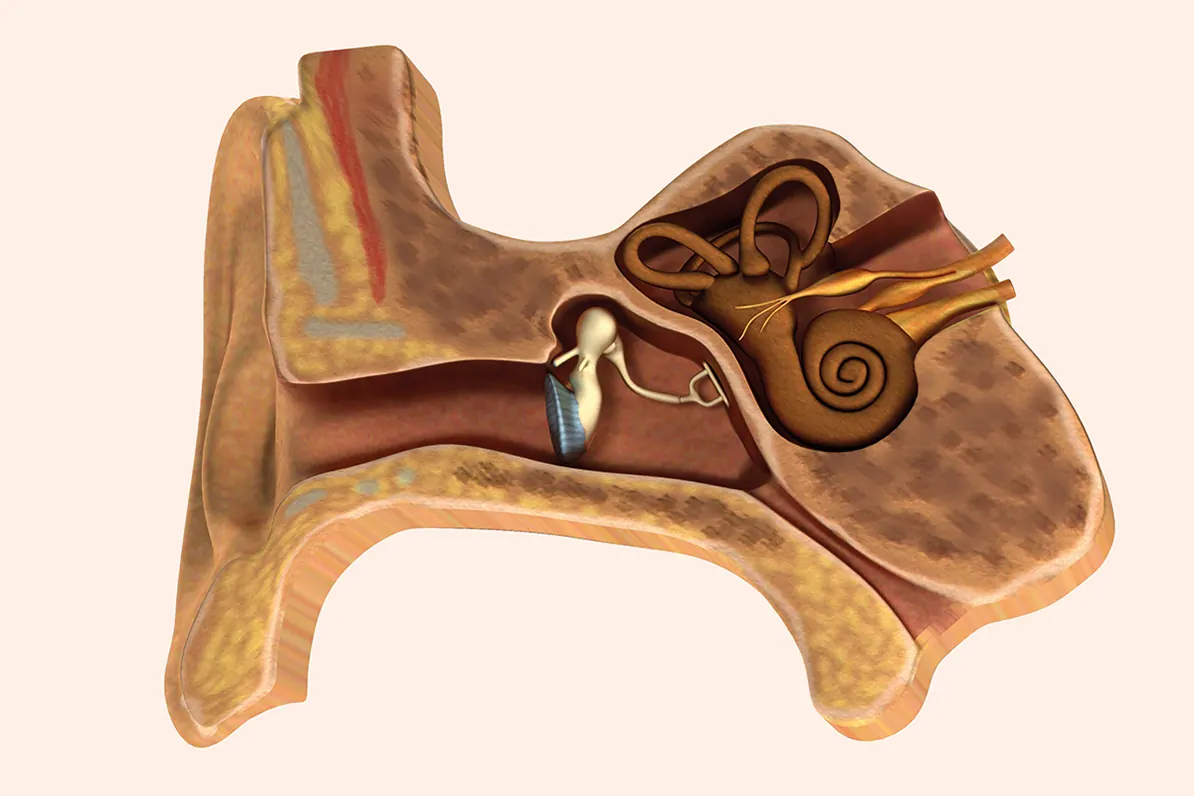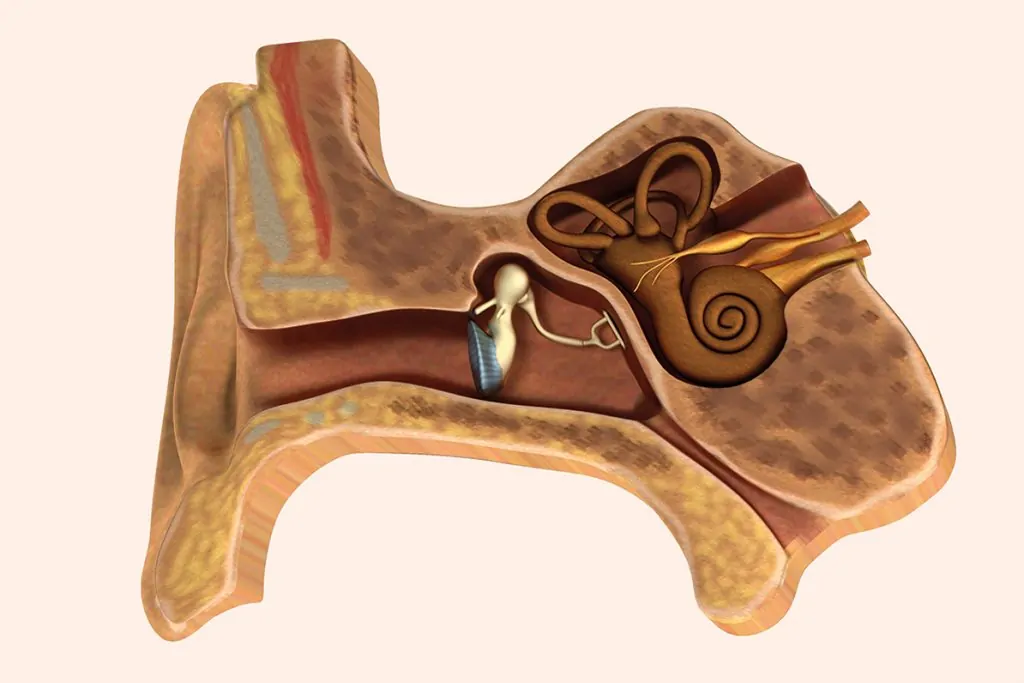
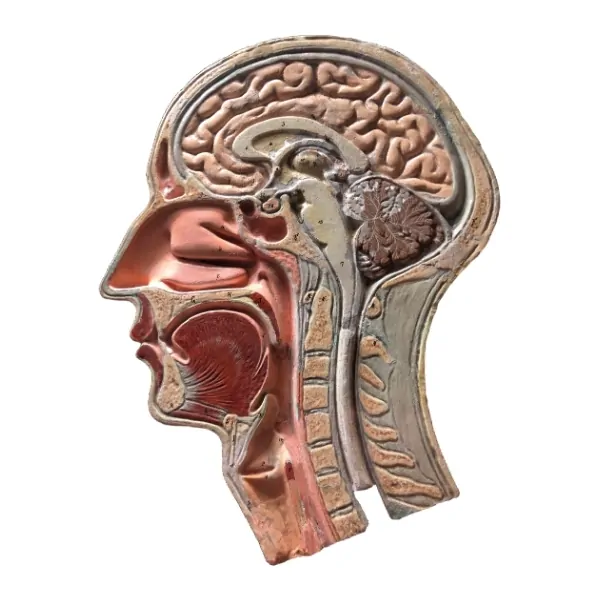
Extraído del libro, Audiología: Teoría y Práctica. Capítulo 2. Anatomía funcional y fisiopatología de los sistemas auditivo, vestibular y fonador.
Origen de los receptores
Desarrollo filogenético
La percepción de la aceleración lineal y angular por los distintos receptores vestibulares permite que todas las especies animales que los poseen puedan orientarse en el espacio terrestre, aéreo y acuático de nuestro planeta. Esencialmente, desde que surgió la función del equilibrio en los primitivos organismos animales prehistóricos ha permanecido sin cambios hasta la actualidad, aunque morfológicamente los órganos sensoriales se han ido especializando y evolucionando según las diversas especies. El más simple es el estatocisto, consistente en una invaginación de la superficie animal (medusa, esponja) con líquido en su interior y una partícula calcárea que hace presión y desplaza los cilios de las células receptoras (localizadas en una región de la pared, similar a la mácula del sáculo).
En función de la fuerza de la gravedad que se ejerce sobre dichas células, estos organismos mantienen una orientación espacial con sentido y dirección vertical. Posteriormente, en algunos moluscos, como el pulpo y la sepia, surgieron las primeras crestas, además del estatocisto, lo que permitió responder a movimientos de aceleración angular, con presencia de nistagmo.
La complejidad del laberinto posterior progresa en un grupo de vertebrados con la aparición de los primeros conductos semicirculares verticales y con el cierre de la invaginación del estatocisto, formando una vesícula aislada en el interior, con líquido de producción endógena (endolinfa). La lamprea alcanza una estructura de canales anterior y posterior (con dilataciones bullosas, las ampollas, cada una con un primitivo receptor en forma de cresta), comunicados por un saco bilobulado con mácula sacular y utricular separadas, donde se localizan las células sensoriales. La aparición del canal semicircular horizontal en los primeros peces óseos y cartilaginosos (con mandíbula) permitió un mayor control del espacio tridimensional. A partir del máximo desarrollo de dichas estructuras vestibulares en los peces modernos (hace 100 millones de años), se ha llegado al más alto grado de perfección morfofuncional del órgano del equilibrio.
En los vertebrados superiores, las vías nerviosas vestibulares centrales son cada vez más complejas debido a un desarrollo paralelo de aquellos sistemas aferentes que intervienen para mantener el equilibrio.
Desarrollo ontogenético
En un embrión humano de 19 a 21 días (2 mm de longitud corono- caudal), en el ectodermo superficial de la porción cefálica a la altura del rombo encéfalo, se diferencian las primitivas células que forman la placoda ótica. Tras su invaginación (fosa ótica), la separación de la superficie dará origen al otocisto o vesícula ótica (28 días). A partir de su porción dorsal derivarán las diferentes partes del sistema vestibular (laberinto posterior) y desde su porción ventral surgirán las estructuras de la cóclea (laberinto anterior).
Hacia la quinta semana (embrión de 8-9 mm) se forman unos pliegues en la pared del otocisto que corresponderán a los receptores vestibulares. Estos se identifican como sáculo, utrículo y los tres conductos semicirculares (a las 6,5 semanas, 14 mm). En la décima semana (50 mm) todo el laberinto membranoso es muy evidente y se forma a su alrededor un modelo cartilaginoso a partir de la cápsula ótica mesenquimal (Sadler, 2012; Suárez y cols., 2007).
Origen de las vías vestibulares centrales
Desarrollo filogenético
En los vertebrados superiores, las vías nerviosas vestibulares centrales son cada vez más complejas debido a un desarrollo paralelo de aquellos sistemas aferentes que intervienen para mantener el equilibrio (visión y propiocepción), cuyas respectivas vías nerviosas interactúan con la vestibular. La organización de los núcleos vestibulares supraespinales, integrados en la formación reticular, se empieza a observar en la lamprea, con dos agrupaciones neuronales (núcleos dorsal y ventral). A partir de los peces teleósteos se identifican cuatro agrupaciones que van aumentando en el número de células en los vertebrados superiores. Las conexiones vestíbulo-espinales son necesarias para el mantenimiento de la orientación corporal en los vertebrados primitivos. Cuando se incorporan funciones más complejas en animales más evolucionados, aparecen conexiones vestíbulo-cerebelosas y vestíbulo-oculares, siendo menos relevantes las vestíbulo-espinales (Bartual y Pérez, 1998).
Desarrollo ontogenético
A partir del primitivo ganglio estatoacústico-facial (embrión humano de 28 días), derivado de la porción ventral del otocisto y alojado en la mesénquima circundante, se diferencia (décima semana) el ganglio espiral (situado cerca del receptor auditivo en la cóclea) y el ganglio vestibular o de Scarpa (próximo al conducto auditivo interno). En estas primitivas neuronas ganglionares van apareciendo unas delgadas prolongaciones citoplasmáticas en polos opuestos de las células. La prolongación periférica (dendrita) se dirige hacia las respectivas regiones del laberinto membranoso, donde se localizarán los órganos sensoriales. La prolongación central (axón) se dirige a regiones del rombo encéfalo donde, a medida que progrese el desarrollo del sistema nervioso central, se diferenciarán las neuronas que constituirán los futuros núcleos vestibulares.
Los órganos sensoriales vestibulares alcanzan una maduración con aspecto semejante al adulto hacia la vigésimo tercera semana de gestación.
Entre la decimoprimera y la decimotercera semana, cuando se empiezan a diferenciar las células sensoriales en los epitelios de las regiones que corresponderán a las máculas y crestas ampulares, también se pueden identificar terminaciones nerviosas aferentes y eferentes, que se distribuyen por dicho epitelio y establecen algunas sinapsis. Los órganos sensoriales vestibulares alcanzan una maduración con aspecto semejante al adulto hacia la vigésimo tercera semana (Bartual y Pérez, 1998; Suárez y cols., 2007).
Malformaciones del sistema vestibular
Las malformaciones del oído interno que afectan a los conductos semicirculares y al acueducto del vestíbulo, son las que suelen causar vértigos en la infancia. Sin embargo, la malformación más frecuente, la dilatación del conducto semicircular horizontal, es raro que se asocie con un trastorno del equilibrio. Los casos de agenesia de los conductos semicirculares son poco frecuentes y suelen ocasionar un trastorno en la marcha.
Las malformaciones del oído interno que afectan a los conductos semicirculares y al acueducto del vestíbulo, son las que suelen causar vértigos en la infancia.
Anatomía del aparato vestibular periférico
Figura 13
Receptores sensoriales del equilibrio
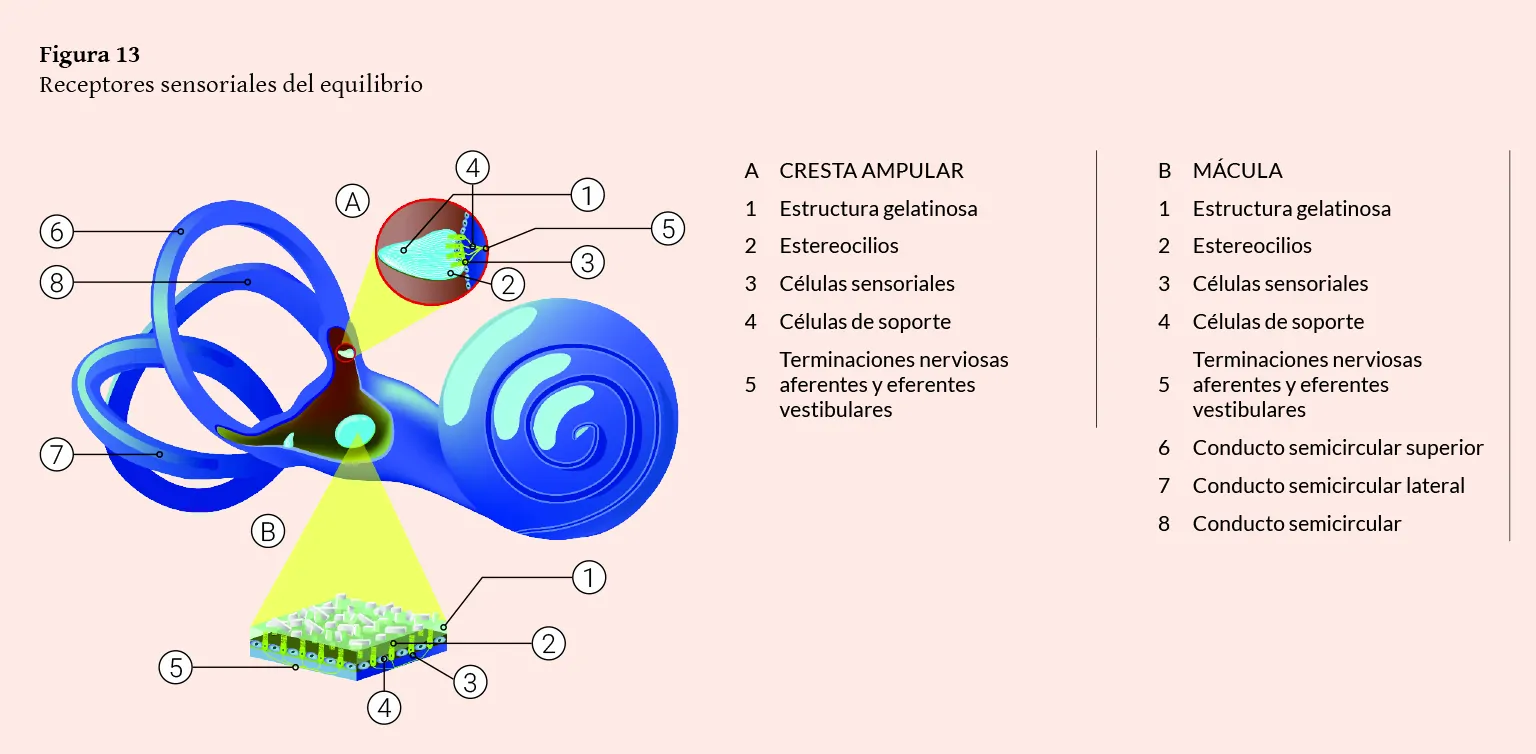
El sistema vestibular está constituido por el aparato vestibular (contenido dentro del oído interno, donde se encuentran los órganos receptores sensoriales periféricos) y por las vías vestibulares o vías nerviosas sensoriales centrales (aferente y eferente).
Vestíbulo
En el interior del vestíbulo del laberinto óseo se distinguen el utrículo y el sáculo del laberinto membranoso. Estos se comunican entre sí por el conducto utrículo-sacular, del que parte el conducto endolinfático (alojado en el acueducto vestibular) que acaba en el saco endolinfático situado en el espacio subdural de la cavidad craneal, al nivel de la cara posterior del peñasco. Las máculas sacular y utricular son órganos receptores integrados por células de soporte y células receptoras sensoriales ciliadas recubiertas por una membrana horizontal, con componentes mucopolisacáridos, sobre la que hay una serie de cristales de carbonato cálcico u otolitos.
En las máculas utricular y sacular existe una línea imaginaria, la estriola, donde se organizan los manojos de células ciliares a ambos lados y con polarizaciones opuestas.
El utrículo es una cavidad conectada a los conductos semicirculares. En el plano horizontal y en su parte anterior, se ubica la mácula (órgano otolítico), pequeña vesícula, aplanada transversalmente y adherida a la fosita semiovoidea, donde se sitúan las células sensoriales o ciliares. Estas son semejantes a las de las ampollas de los conductos semicirculares (con estereocilios y un kinocilio) y con la misma actividad eléctrica. La mácula del utrículo, al estar colocada en el suelo, tiene una orientación horizontal, captando las lateralizaciones hacia los lados, o las inclinaciones de la cabeza y sus desplazamientos lineales hacia atrás y hacia delante. El sáculo está situado por debajo del utrículo, es una pequeña vesícula redondeada adherida a la fosita hemisférica. Al nivel de esta fosita se encuentra la mácula del sáculo.
En las máculas utricular y sacular existe una línea imaginaria (estriola) donde se organizan los manojos de células ciliares a ambos lados y con polarizaciones opuestas. Los estereocilios, están inmersos en una sustancia gelatinosa, la membrana otolítica, que soporta concreciones calcáreas (carbonato cálcico), los otolitos o estatoconias. Estos ejercen una acción gravitacional sobre el conjunto de estereocilios y de la sustancia gelatinosa. Los otolitos están anclados en la masa gelatinosa mediante fibras de colágeno, pero pueden desprenderse y disolverse por el espacio endolinfático (Bartual y Pérez, 1998; Suárez y cols., 2007; Williams, 1998).
Conductos semicirculares
En el interior de los tres conductos semicirculares óseos se encuentran los membranosos, que comunican con el utrículo alojado en el vestíbulo óseo. Están dispuestos en ángulo recto uno respecto al otro, en los tres planos del espacio: los dos de posición vertical son los conductos semicirculares anterior y posterior, y el horizontal, es el conducto semicircular lateral. Tal posición hace posible que detecten la aceleración o desaceleración rotacional. Cada uno presenta una dilatación en su parte inferior denominada ampolla, en el interior de la cual se encuentra una protrusión con las células del órgano receptor del equilibrio o cresta ampular. Cada cresta contiene un grupo de células sensoriales ciliadas y de sostén cubiertas por una pequeña masa de material gelatinoso, la cúpula. De esta forma se crea un espacio hermético que ocluye la luz del conducto impidiendo la circulación libre de la endolinfa. Hay dos tipos de células sensoriales en las crestas ampulares: las células tipo I, que son las que tienen apariencia de botellón adelgazadas en su parte superior, donde, en el borde libre, se hallan los estereocilios de distintos grosores que se dirigen hacia el espacio endolinfático; y las células tipo II, que son de forma cilíndrica y también tienen estereocilios.
El nervio vestibular conducirá la información hasta el interior del cerebro, uniéndose al nervio coclear para constituir el VIII par craneal o nervio estatoacústico o cócleo-vestibular, que atraviesa el conducto auditivo interno de la porción petrosa del hueso temporal.
Existe un cilio más grueso que los demás, el kinocilio o cinocilio, que está ubicado en uno de los extremos de la célula. Los otros estereocilios se sitúan al lado. Los estereocilios de cada célula ciliar se disponen en cinco hileras decrecientes de ocho cilios cada una. Cada estereocilio posee una porción intracelular y otra mayor extracelular, continuación una de la otra, pudiendo apreciarse en estos una fina estructura o fibrilla centrales y una porción periférica a modo de vaina de aquella. Por tanto, cada célula posee por término medio unos cuarenta estereocilios y un kinocilio. La orientación de este depende del conducto; así, en el conducto lateral, el kinocilio está ubicado en el lado utricular, y en los conductos anterior y posterior, en el sentido opuesto al utrículo.
Al mover la cabeza, se desplazan con ella los conductos semicirculares membranosos y las células ciliadas, desencadenando el fenómeno de transducción, es decir, la transformación de la energía mecánica en corriente eléctrica de tipo nervioso. En la base de las células sensoriales establecen sinapsis las prolongaciones de las neuronas del ganglio vestibular de Gasser. De ellas se originan los axones que forman el nervio vestibular, el cual conducirá la información hasta el interior del cerebro, uniéndose al nervio coclear para constituir el VIII par craneal o nervio estatoacústico o cócleo-vestibular, que atraviesa el conducto auditivo interno de la porción petrosa del hueso temporal (Bartual y Pérez, 1998; Suárez y cols., 2007; Williams, 1998).
Funciones del sistema vestibular
Para comprender los mecanismos fisiológicos del sistema vestibular se definen los siguientes conceptos básicos:
– Equilibrio: estado de un cuerpo sometido a dos o varias fuerzas cuya resultante es nula, permitiendo que el cuerpo permanezca estable.
– Desequilibrio: perturbación del equilibrio que aparece mientras se permanece de pie o durante la marcha. Si se pierde el control postural, se produce la caída.
– Orientación espacial: capacidad del individuo para orientarse en relación al medio que le rodea.
El sentido del equilibrio es un sentido fisiológico que permite al individuo mantener una postura respecto a la fuerza de la gravedad, la orientación espacial, la estabilización de la imagen en la retina y coordinación de respuestas motoras.
– Fuerza de la gravedad: es la referencia fundamental para el mantenimiento del equilibrio y para la orientación espacial.
– Vertical subjetivo: el equilibrio y la orientación dependen de este sentido que está presente en todo individuo. Gracias a ella, se permite el mantenimiento del equilibrio en posición vertical tomando como referencia la gravedad.
– Actitud: postura propia de una especie animal característica de su modo de locomoción. En el humano sería la postura eréctil, que es una posición frágil y difícil de mantener, tanto en situaciones estáticas como dinámicas o de movimiento lineal y angular (rotatorio).
El sentido del equilibrio es un sentido fisiológico que permite al individuo mantener una postura respecto a la fuerza de la gravedad y la orientación espacial de su cuerpo, además de estabilizar la imagen en la retina, especialmente durante la marcha y coordinar respuestas motoras. Existen cuatro sistemas sensoriales que mandan información de equilibrio al sistema nervioso central: el sistema vestibular, el propioceptivo, el visual y el táctil.
Las máculas del utrículo y sáculo del vestíbulo informan de las posiciones de la cabeza en su relación con la gravedad y con la aceleración de tipo lineal. Los conductos semicirculares advierten de la aceleración angular. El sistema propioceptivo está integrado por receptores sensoriales músculo-tendinosos que reportan a los centros nerviosos superiores la posición de las articulaciones, tendones, músculos, etc. El tacto contribuye informando de los puntos de contacto de la superficie corporal con el entorno. El sistema visual tiene una gran importancia ya que existen abundantes conexiones nerviosas con el sistema vestibular para informar de la posición del cuerpo y ayudar a mantener el equilibrio.
La salida efectora es el sistema motor. Con la información que llega al cerebro del sistema vestibular, más la entrada visual, táctil y cinestésica o propioceptiva (fundamentalmente de los músculos de la nuca y miembros inferiores) se puede determinar la orientación motora del cuerpo en el espacio. Además, el cerebro es capaz de conocer la relación de cada uno de sus miembros entre sí y mantener el equilibrio y la postura erecta durante la marcha.
A pesar de que el sistema vestibular ha sido diseñado para detectar los movimientos cefálicos en todas las posibles direcciones del espacio, hay limitaciones. Estas consisten en la incapacidad de detectar movimientos lentos de la cabeza a velocidades constantes en el plano horizontal y en la imposibilidad de distinguir la inclinación de la cabeza en una aceleración lineal.
FUNCIÓN DE LOS ÓRGANOS SENSORIALES DEL APARATO VESTIBULAR
En los mecanorreceptores ampulares de los conductos semicirculares no se han encontrado diferencias funcionales entre sus dos tipos de células. Ambas, en ausencia de movimiento ciliar, provocan una descarga eléctrica basal.
Cuando hay desplazamiento de la endolinfa y se movilizan los estereocilios hacia el kinocilio, la descarga basal aumenta (excitación); si tiene una dirección contraria, disminuye (inhibición). Ante un movimiento de la cabeza hacia la izquierda, en el conducto lateral o externo izquierdo habrá un desplazamiento ampulípeto (hacia la ampolla, por la inercia de la endolinfa) con desviación de la cresta hacia el utrículo, mientras que en el derecho habrá un movimiento inverso (ampulífugo).
A consecuencia de esto, habrá un aumento de la descarga basal en el lado izquierdo y una disminución en el derecho. Por lo tanto, cuando hay un movimiento de la cabeza, hay un aparato sensorial que aumenta sus descargas y otro que las disminuye. Si sigue la rotación se detiene la activación, dado que la endolinfa y la cresta igualan su desplazamiento con el de la cabeza. Cuando cesa, el proceso se invierte y vuelven a activarse los órganos sensores. De esta forma, los canales semicirculares informan del inicio y del final de la rotación, y no de la rotación en sí. En los otros planos cualquier movimiento angular estimulará como mínimo un par de canales semicirculares.
Con la información que llega al cerebro del sistema vestibular, más la entrada visual, táctil y cinestésica o propioceptiva se puede determinar la orientación motora del cuerpo en el espacio.

Los conductos semicirculares predicen de antemano un desequilibrio. Cuando se produce un movimiento rotatorio que produce desequilibrio se activan dos canales semicirculares, mientras que los otros cuatro actúan de manera opuesta para así favorecer la vuelta a una posición de equilibrio nueva.
Aunque con menos efectividad que las aceleraciones y desaceleraciones lineales, las máculas son también estimuladas por las fuerzas gravitatorias y por las aceleraciones de traslación centrífugas y centrípetas.
Para entender la función de los órganos sensoriales ampulares se toma como ejemplo el canal semicircular horizontal. Además, estos son los más importantes fisiológicamente ya que son estimulados con la aceleración angular de la cabeza en el plano horizontal (movimiento de negar con la cabeza). La estimulación de un canal semicircular horizontal da lugar a la activación de diferentes grupos musculares del cuerpo: musculatura ocular (contracción del músculo recto interno homolateral y externo contralateral), musculatura axial (contracción músculos homolaterales del tronco) y musculatura de las extremidades (contracción músculos extensores homolaterales y flexores contralaterales). Por ello, en reposo, la actividad de ambos conductos semicirculares horizontales es equivalente y hay un reparto uniforme del tono muscular en todo el cuerpo.
El nistagmo se caracteriza por movimientos alternantes oculares. Consta de dos fases: una rápida y otra lenta. El sentido del nistagmo viene dado por el movimiento que se realiza en la fase rápida. Un movimiento muy amplio de la cabeza hacia la izquierda originaría un movimiento ocular hacia la derecha para compensar y poder tener una visión correcta. Sin embargo, si la amplitud del movimiento es muy grande, el ojo no va a girarse sobre sí mismo, por ello vuelve a su posición normal en un rápido movimiento hacia la izquierda.
La vía vestibular aferente comprende el conjunto de neuronas conectadas entre sí y en sinapsis con las células de los receptores del aparato vestibular para transmitir señales nerviosas al tronco del encéfalo y a la corteza cerebral.
Las leyes de Ewald fueron establecidas para explicar la fisiología del laberinto posterior y se enuncian así:
1ª Ley: el movimiento de la endolinfa es el mismo que el de la desviación del cuerpo y extremidades y que la fase lenta del nistagmo.
2ª Ley: en el canal horizontal, la excitación que da el movimiento ampulípeto es mayor que la del ampulífugo. En el canal vertical es al contrario.
3ª Ley: en los canales semicirculares verticales se invierte la segunda ley, es decir, es más activa la corriente ampulífuga que la ampulípeta (Bartual y Pérez, 1998; Haines, 2013; Purves, 2015).
Anatomía funcional de las vías vestibulares centrales
Vías vestibulares aferente y eferente
Desde cada uno de los receptores vestibulares (tres crestas ampulares y dos máculas) se identifican fibras aferentes que constituyen fascículos, los cuales transmiten información independiente hasta llegar al sistema nervioso central. No obstante, se agrupan para formar dos divisiones del nervio vestibular a su entrada al conducto auditivo interno. La división superior está formada por fibras utriculares, algunas saculares y las de los conductos semicirculares horizontal y anterior. La división inferior del nervio vestibular se constituye por la mayor parte de las fibras saculares y las del canal posterior.
La porción ventral del núcleo vestibular lateral o de Deiters proyecta al núcleo del motor ocular común e interviene en la producción de los reflejos vestíbulo-oculares.
En el conducto auditivo interno, por delante y por debajo del nervio vestibular, se dispone el nervio coclear o auditivo formado por fibras provenientes de la cóclea. Ambos nervios se unifican anatómicamente en el VIII par craneal o nervio vestíbulococlear o estatoacústico. Junto a las fibras aferentes, se encuentran fibras eferentes originadas en el sistema nervioso central para realizar un control o modulación de la actividad de los órganos receptores periféricos (Bartual y Pérez, 1998; Suárez y cols., 2007).
La vía vestibular aferente comprende el conjunto de neuronas conectadas entre sí que, partiendo de sinapsis con las células sensoriales de los receptores del aparato vestibular (dos máculas y tres crestas ampulares), transmiten señales nerviosas al tronco del encéfalo y a la corteza cerebral haciendo escala en otras regiones encefálicas. A lo largo de todo este trayecto hay asociaciones con la información recibida por los sistemas sensoriales visual y propioceptivo.
El nervio vestibular (raíz vestibular del VIII par craneal) surge de las neuronas bipolares en el ganglio vestibular o ganglio de Scarpa, primer nivel de la vía vestibular aferente. Las fibras periféricas o dendritas terminan en las células sensoriales de las crestas ampulares de los conductos semicirculares y máculas del sáculo y del utrículo. Las fibras centrales o axones entran lateralmente en el bulbo raquídeo y pasan entre el pedúnculo inferior y el tracto espinal del trigémino. Estas fibras aferentes se bifurcan en ramas ascendentes y descendentes y hacen sinapsis con neuronas situadas en los núcleos vestibulares, segundo nivel de la vía vestibular aferente.
Los núcleos vestibulares están formados por cuatro áreas principales situadas lateralmente y debajo del suelo del cuarto ventrículo. Las proyecciones de estos núcleos van por el fascículo longitudinal medial, cordón nervioso largo y delgado que corre a ambos lados de la línea media que va hacia abajo (bulbo y médula espinal), y hacia arriba (a los lados del acueducto de Silvio) y termina en los núcleos del III par, conectando así los núcleos vestibulares con los núcleos motores del ojo, cuello, extremidades y tronco (Bartual y Pérez, 1998; Haines, 2013; Purves, 2015; Suárez y cols., 2007).
El fastigius es regulado por el pyramis, que a su vez está controlado por la corteza cerebral a través de la vía córticopontocerebelosa.
El núcleo vestibular superior o de Betcherew, en posición rostral, recibe las aferencias de las crestas ampulares. De aquí salen proyecciones (fascículo longitudinal medial) a las neuronas motoras del núcleo troclear (patético o IV par craneal) homolateral y al núcleo motor ocular común homolateral y contralateral. Está implicado en el reflejo vestíbulo-oculomotor en el plano vertical.
El núcleo vestibular lateral o de Deiters recibe colaterales de las crestas ampulares en su porción dorsal y proyecciones de las máculas en porción ventral. La porción dorsal da lugar al tracto vestíbulo
-espinal lateral que proyecta contralateralmente a las interneuronas y motoneuronas que inervan de forma inhibitoria a los músculos extensores de las extremidades. Contribuye al mantenimiento del tono de los músculos antigravitatorios. La porción ventral de este núcleo proyecta al núcleo del motor ocular común e interviene en la producción de los reflejos vestíbulo-oculares.
El núcleo vestibular medial o principal (Schwalbe) recibe aferencias de las máculas y colaterales que llegan al núcleo lateral. Su proyección va por el fascículo longitudinal medial, ascienden y descienden directas y cruzadas; las ascendentes constituyen la vía oculomotora y sinaptan con las motoneuronas y neuronas del núcleo motor ocular externo homolateral y contralateral (responsable del nistagmo horizontal) y con las del núcleo motor ocular común. También emite fibras para los núcleos motores del cuello y centros vegetativos. Las fibras desencadenantes forman la vía vestibulo-espinal.
Además, envía fibras hacia la sustancia reticular media y núcleos del neumogástrico siendo responsables de reflejos vegetativos como náuseas, vómitos, sudoración palidez, diarrea, etc. Participa en los reflejos vestíbulo-oculares, sobre todo, los horizontales y en los reflejos posturales compensatorios.
El núcleo vestibular inferior o espinal (Roller) recibe aferencias periféricas del sáculo y utrículo y algunas fibras colaterales de las crestas. Su salida forma parte de las vías vestíbulo-espinales e integra las señales periféricas con las llegadas del cerebelo, teniendo un efecto inhibitorio sobre los músculos extensores contralaterales. La acción que ejerce sobre los reflejos y tono muscular se hace a través de este.
Relación vestibular-cerebelar y cerebelar-vestibular: la mayoría de las aferencias vestibulares también se dirigen homolateralmente a través del cuerpo restiforme hacia los núcleos floconodular y fastigius del cerebelo. De estos se envían terminaciones a los núcleos vestibulares medial e inferior, inhibiendo a las neuronas vestibulares de segundo orden. Del fastigius salen fibras cruzadas que inhiben el área vestibular contralateral. El fastigius es regulado por el pyramis, que a su vez está controlado por la corteza cerebral a través de la vía corticopontocerebelosa.
La función principal del cerebelo es el mantenimiento del equilibrio por medio del núcleo fastigius y lóbulo floconodular, y su lesión da lugar a trastornos: en reposo, caída hacia adelante, atrás o lateral, y en movimiento, marcha bamboleante con las piernas separadas para aumentar la base de sustentación. El cerebelo también regula el tono muscular y coordina los movimientos de todo el cuerpo.
A la corteza cerebral también llegan proyecciones de los núcleos vestibulares, específicamente al lóbulo temporal (circunvolución temporal superior) a través de la vía reticulotalámica.
Las vías eferentes vestibulares están constituidas por el fascículo eferente de Petroff y Gacek. Sus fibras se distribuyen por las células sensoriales a razón de una fibra eferente por cada 60 fibras aferentes (Suárez y cols., 2007).
Reflejos vestibulares
Los cambios espaciales estimulan los reflejos posturales vestibulares, ayudando a mantener el equilibrio, la postura y la mirada. De esta forma se anticipa al desequilibrio que se generará en cuestión de segundos ante un desplazamiento del cuerpo en el espacio (Suárez y cols., 2007).
REFLEJO VESTÍBULO-OCULAR
Desempeña una importante función, cuando se cambia de forma brusca de posición o incluso con el movimiento de la cabeza, permitiendo mantener estable la mirada en la retina.
Este reflejo se puede observar también en personas ciegas. Cada vez que la cabeza rota en una dirección los ojos rotan suavemente en la dirección opuesta.
El reflejo actúa, por ejemplo, al producirse un movimiento hacia la izquierda. La endolinfa se desplaza dentro de los canales semicirculares hacia el lado opuesto, aumenta la descarga hacia los núcleos vestibulares de aquí, y las fibras que van a los núcleos óculomotores aumentan la actividad del músculo recto lateral derecho del ojo e inhiben al músculo recto medial.
Los cambios espaciales estimulan los reflejos posturales vestibulares, ayudando a mantener el equilibrio, la postura y la mirada.
Cuando el giro se interrumpe, los ojos siguen moviéndose en la dirección contraria y después vuelven rápidamente a la posición de la línea media con un movimiento de sacudida (nistagmo vestibular) (Suárez y cols., 2007).
REFLEJOS POSTURALES Y DEL EQUILIBRIO
La orientación espacial está basada en la interacción visual, vestibular y cinestésica que permite la coordinación de los movimientos. La vía para los reflejos del equilibrio comienza en los nervios vestibulares, que pasan cerca del cerebelo y de los núcleos vestibulares desde donde se envían señales hacia los núcleos reticulares. Hay señales que van hacia la médula espinal y el cerebelo adapta el tono muscular para cubrir la nueva situación.
La función principal del cerebelo es el mantenimiento del equilibrio y su lesión da lugar a trastornos como la caída hacia adelante, atrás o lateral estando en reposo, o la marcha bamboleante, en movimiento.
El área vestíbulo-cerebelosa, es importante en el control del equilibrio, sobre todo en la ejecución de movimientos rápidos. Calcula, a partir de distintas velocidades y direcciones, dónde estarán las distintas partes del cuerpo en los próximos milisegundos.
Durante los cambios de posición, el sistema vestibular tiene una influencia estimuladora en el control autonómico respiratorio, modificándose esta actividad muscular.
Los estímulos vestibulares asociados a movimientos de la cabeza realizan un rol inhibitorio vagal mediante el control del reflejo barorreceptor (Suárez y cols., 2007).